


222-223
Организация клетки. Биомембраны
Транспортные белки
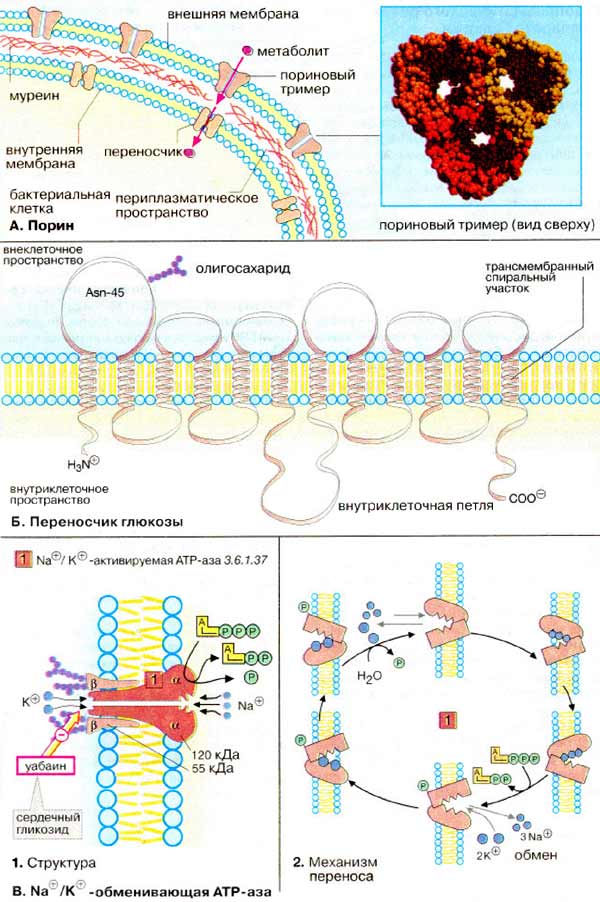 А. Порин
А. Порин
Бактерии имеют одну, но достаточно
сложно устроенную клеточную стенку. Плазматическая мембрана грамотрицательных
бактерий защищена от внешней среды сетью пептидогликанов (муреин, см. с. 46) и
дополнительной наружной мембраной. Метаболиты, которые бактериальной
клетке необходимо абсорбировать или высвободить, должны иметь возможность без
труда пересекать наружную мембрану. Для обеспечения процесса переноса у бактерий
имеются трансмембранные каналообразующие белки, так называемые
порины. Эти белки - тримеры образуют поры, заполненные водой и
проницаемые для молекул с молекулярной массой до 600 Да (облегченная или
опосредованная диффузия). В высших организмах пориноподобные белки найдены в
мембранах митохондрий и хлоропластов.
На рисунке справа показана структура
тримера порина, находящегося в наружной мембране бактерии Rhodopseudomonas
blastica.
Б. Переносчик
глюкозы
Как описано на с. 220, захват глюкозы
клетками обычно является опосредованным процессом. Сначала глюкоза связывается с
переносчиком глюкозы [ГЛУТ (GLUT)], локализованным в клеточной мембране.
Перенос глюкозы через мембрану обеспечивается за счет изменения конформации
молекулы переносчика.
Для многочисленных мембранных белков известна лишь последовательность
аминокислот, но не трехмерная структура. Из-за трудностей кристаллизации мембранных
белков к настоящему времени известны трехмерные структуры только некоторых из
них (см., например, схему А и с. 216).
Здесь приведена структура переносчика глюкозы, содержащего 12 трансмембранных
α-спиральных фрагментов (см. с. 216) и один олигосахарид,
ориентированный в окружающую среду.
Переносчики глюкозы представляют собой
семейство структурно близких мембранных белков о различными функциями. ГЛУТ-1 и
ГЛУТ-3 имеют высокое сродство к глюкозе (Кd около 1 мМ). Они
обнаружены почти во всех клетках, где обеспечивают постоянное поступление
глюкозы. ГЛУТ-2 найден в клетках печени и поджелудочной железы. Этот переносчик
обладает гораздо меньшим сродством к глюкозе (Kd 15-20 мМ). Это
означает, что связывание глюкозы ГЛУТ-2 пропорционально концентрации глюкозы в
крови. ГЛУТ-4 с Kd около 5 мМ найден в плазматической мембране
мышечных и жировых клеток. Гормон инсулин вызывает увеличение количества молекул
ГЛУТ-4 на поверхности клетки и таким образом стимулирует поступление глюкозы в
эти ткани. ГЛУТ-5 синтезируется клетками кишечного эпителия. Этот переносчик
обеспечивает симпорт глюкозы с ионами Na+ (см. с.
220).
В. Na+/K+
-обменивающие АТФ-азы
АТФ-азные (АТР-азные) системы,
транспортирующие ионы К+ и Na+ относятся к группе
транспортных белков. Они осуществляют АТФ-зависимый активный транспорт
через мембраны против концентрационного градиента. Имеется множество различных
АТФ-аз, способных транспортировать различные вещества от неорганических катионов
(ионные насосы) до пептидов (пептидные насосы) и неполярных соединений
(например, переносчики лекарственных веществ или белки, обеспечивающие
множественную лекарственную устойчивость). Во всех клетках имеются ионные насосы
(ионтранспортирующие АТФ-азы), осуществляющие постоянный перенос таких катионов,
как Н+ и Na+, K+ и Са2+, что
существенно важно для поддержания электрохимического градиента (см. с.
220).
На схеме показана
Na+/K+-обменивающая АТФ-аза, которая найдена в
плазматической мембране практически всех животных клеток. Это мембранный
гликопротеин, состоящий из четырех субъединиц (α2β2).
Цитоплазматическая область фермента участвует в реакционном цикле
фосфорилирования/дефосфорилирования, принимая попеременно два конформационных
состояния, ответственных за транспорт ионов. При расходовании одной молекулы АТР
из клетки «выкачивается» три иона Na+ в обмен на поступающие два иона
К+. Благодаря непрерывному функционированию этого «насоса»
обеспечивается поддержание неравновесного распределения ионов Na+ и
К+ между цитоплазмой клетки и окружающей средой, характерное для
животных клеток (см. с. 340).