


116-117
Метаболизм. Регуляция
Механизмы регуляции метаболических процессов
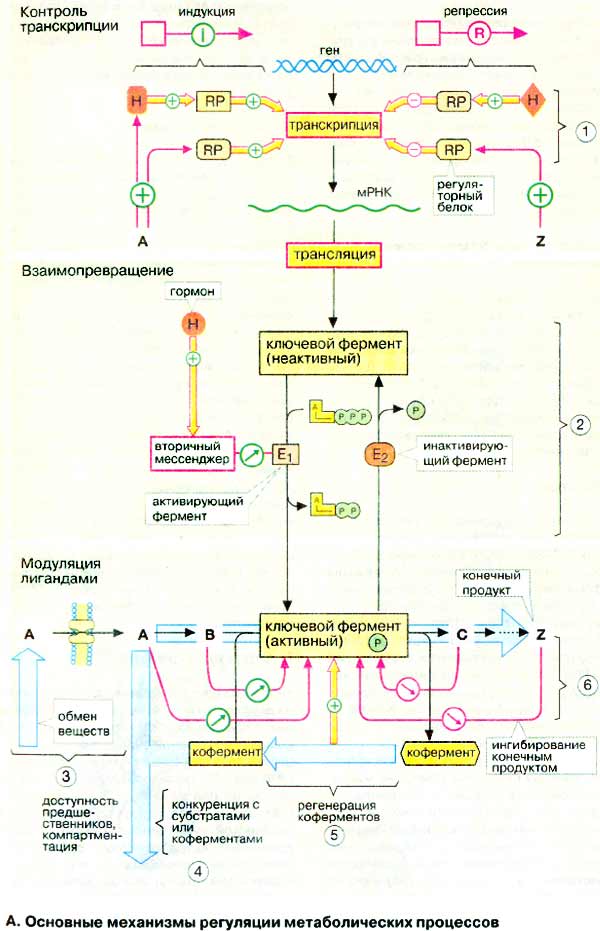 А. Основные механизмы регуляции
метаболических процессов
А. Основные механизмы регуляции
метаболических процессов
Активность всех путей обмена веществ постоянно регулируется,
что обеспечивает соответствие синтеза и деградации метаболитов физиологическим
потребностям организма. В этом разделе рассматриваются механизмы такой регуляции.
Более детально вопросы регуляции клеточного метаболизма представлены на сс.
118-123.
Поток метаболитов в обмене веществ
определяется прежде всего активностью ферментов (см. с. 94). Для
воздействия на тот или иной путь достаточно регулировать активность фермента,
катализирующего наиболее медленную стадию. Такие ферменты, называемые
ключевыми ферментами, имеются в большинстве метаболических путей.
Активность ключевого фермента регулируется на трех независимых
уровнях,
Контроль транскрипции. Контроль за биосинтезом фермента
(1) осуществляется на генетическом уровне. Прежде
всего речь идет о синтезе соответствующей мРНК (mRNA), а также о транскрипции
кодирующего фермент гена, т.е. о регуляции транскрипции (см. с. 120,
242). В этом процессе принимают участие регуляторные
белки (RP) (факторы транскрипции), действие которых направлено непосредственно
на ДНК. К тому же в генах имеются специальные регуляторные участки — промоторы
— и участки связывания регуляторных белков (регуляторные элементы). На эффективность
действия этих белков влияют метаболиты или гормоны. Если этот механизм усиливает
синтез фермента, говорят об индукции, если же снижает или подавляет —
о репрессии. Процессы индукции и репрессии осуществляются лишь в определенный
отрезок времени.
Взаимопревращение. Значительно
быстрее, чем контроль транскрипции, действует взаимопревращение ключевых
ферментов (2). В этом случае фермент присутствует в клетке в неактивной форме.
При метаболической потребности по сигналу извне и при посредничестве вторичного
мессенджера активирующий фермент
(E1) переводит ключевой фермент в каталитически активную форму.
Если потребность в этом пути обмена веществ отпадает, инактивирующий
фермент (E2) снова переводит ключевой фермент в неактивную форму.
Процесс взаимопревращения в большинстве случаев состоит в АТФ-зависимом
фосфорилировании ферментных белков протеинкиназой и соответственно
дефосфорилировании фосфатазой (см. с. 122). В большинстве случаев более
активна фосфорилированная форма фермента, однако встречаются также и
противоположные случаи.
Модуляция лигандами. Важным
параметром, контролирующим протекание метаболического пути, является потребность
в первом реагенте (здесь это метаболит А). Доступность метаболита А возрастает с
повышением активности метаболического пути (3), в котором образуется А, и падает
с повышением активности других путей (4), в которых А расходуется. Доступность А
может быть ограничена в связи с его транспортом в другие отделы
клетки.
Часто лимитирующим фактором является
также доступность кофермента (5). Если кофермент регенерируется по
второму независимому пути, этот путь может лимитировать скорость основной
реакции. Таким образом, например, гликолиз и цитратный цикл регулируются
доступностью НАД+ (см. с. 148). Так как НАД+
регенерируется в дыхательной цепи, последняя регулирует катаболизм глюкозы и
жирных кислот (контроль дыхания, см. с. 146).
Наконец, активность ключевого фермента может регулироваться
лигандом (субстратом, конечным продуктом реакции, коферментом, другим
эффектором) как аллостерическим эффектором путем связывания его не в
самом активном центре, а в другом месте фермента, и вследствие этого изменением
ферментативной активности (6, см. с. 118).
Ингибирование ключевого фермента часто вызывается конечными продуктами реакции
соответствующей метаболической цепи (ингибирование по типу обратной связи)
или метаболитом, участвующим в другом пути. Стимулировать активацию фермента
может также первый реагент реакционной цепи.